Difference between revisions of "Schizodactylus groeningae"
m (Using Gallery Template for all images) |
m (Fixing citation) |
||
| (2 intermediate revisions by one other user not shown) | |||
| Line 8: | Line 8: | ||
| title = On the placement of the Cretaceous orthopteran Brauckmannia groeningae from Brazil, with notes on the relationships of Schizodactylidae (Orthoptera, Ensifera) | | title = On the placement of the Cretaceous orthopteran Brauckmannia groeningae from Brazil, with notes on the relationships of Schizodactylidae (Orthoptera, Ensifera) | ||
| journal = ZooKeys | | journal = ZooKeys | ||
| − | |||
| volume = 77 | | volume = 77 | ||
| pages = 17--30 | | pages = 17--30 | ||
| doi = 10.3897/zookeys.77.769 | | doi = 10.3897/zookeys.77.769 | ||
| + | | citationurl = http://www.pensoft.net/journal_home_page.php?journal_id=1&page=article&type=citation&article_id=769 | ||
| + | | publisher = Pensoft Publishers | ||
| + | | publisherurl = http://www.pensoft.net/ | ||
}} | }} | ||
| − | |||
==Taxonavigation== | ==Taxonavigation== | ||
| Line 22: | Line 23: | ||
==Name== | ==Name== | ||
| − | * ''[[Schizodactylus groeningae]]'' ({{aut|Martins-Neto}}, 2007) | + | * ''[[Schizodactylus groeningae]]'' ({{aut|Martins-Neto}}, 2007) {{External databases}} |
| − | ''[[Brauckmannia groeningae]]'' Martins-Neto 2007: 4, fig 1. | + | ''[[Brauckmannia groeningae]]'' Martins-Neto 2007: 4, fig 1. |
| − | + | ||
==Material examined== | ==Material examined== | ||
Latest revision as of 16:55, 15 March 2011
| Notice: | This page is derived from the original publication listed below, whose author(s) should always be credited. Further contributors may edit and improve the content of this page and, consequently, need to be credited as well (see page history). Any assessment of factual correctness requires a careful review of the original article as well as of subsequent contributions.
If you are uncertain whether your planned contribution is correct or not, we suggest that you use the associated discussion page instead of editing the page directly. This page should be cited as follows (rationale):
Citation formats to copy and paste
BibTeX: @article{Heads2011ZooKeys77, RIS/ Endnote: TY - JOUR Wikipedia/ Citizendium: <ref name="Heads2011ZooKeys77">{{Citation See also the citation download page at the journal. |
Ordo: Orthoptera
Familia: Schizodactylidae
Genus: Schizodactylus
Name
- Schizodactylus groeningae (Martins-Neto, 2007) Pensoft Taxon Profile {{#widget:TaxonProfile
|id=Schizodactylus groeningae |height=500
}}
Brauckmannia groeningae Martins-Neto 2007: 4, fig 1.
Material examined
Near-complete adult (sex indet.), MfNB-I.2079. Brazil, Ceará, Chapada do Araripe; Crato Formation, Nova Olinda Member, Lower Cretaceous.
Diagnosis
Schizodactylus groeningae is distinguished from all congeners by the following characters: [1] robust and acutely subovoid metatibiae (in all extant species the metatibiae are of equal width along their entire length); and [2] distinctive blade- or paddle-like lateral processes arising in the distal half of the metabasitarsus (in all extant species these processes are triangular with a broad base and acute, posteriorly directed apex, and arise within the proximal half of the metabasitarsus).
Description of MfNB-I.2079
Large, near-complete specimen preserved in slightly oblique, dorsal aspect (Figs 1, 2). Head capsule robust, 8.46 mm wide at genae; vertex short, c. 2.43 mm from occipital margin as preserved; occipital foramen large, broad; interocular distance 3.98 mm; compound eyes large, c. 2.61 mm wide dorsally. Pronotum markedly wider than long with distinctive marginal sulci; medial length 3.41 mm; width 9.36 mm. Pterothorax poorly preserved, c. 9.20 mm long. Wings incompletely preserved basally, extending posteriorly beyond abdominal apex, tightly folded in a distinctive apical coil. Abdomen somewhat crushed dorsolaterally, c. 16.12 mm long as preserved (apical part missing); first tergite (tg1) largely indistinct, at least 1.45 mm long; tg2 1.96 mm long; tg3 1.83 mm long; tg4 2.17 mm long; tg5 2.35 mm long; remaining tergites incompletely preserved but shorter than previous tergites; right lateral parts of abdominal sternites 1 through 5 visible next to corresponding tergal sclerites; pleural margin distinct. Total body length measured from fastigium verticis to abdominal apex 34.91 mm. Profemora robust and laterally compressed, 12.33 mm long; left profemur preserved in dorsal aspect, with distinct longitudinal dorsal carina; right profemur preserved in lateral aspect, with prominent transverse dorsolateral striae and distinct longitudinal inferior carina. Protibiae robust and lateral compressed, markedly inflated and acutely subovoid in form, 10.24 mm long; left protibia incompletely preserved in oblique dorsal aspect, bearing at least three spur sockets on outer lateral margin, spurs themselves not preserved; right protibia preserved in lateral aspect and somewhat crushed. Left prothoracic leg (L1 in Fig. 2) with distal part missing, only apical part of ungues preserved. Right prothoracic leg (R1 in Fig. 2) with tarsus incompletely preserved, at least 6.52 mm in total length; basitarsus subcylindrical in form, inflated apically, at least 1.75 mm long; second tarsomere short, c. 1.21 mm long, with prominent blade-like lateral process, 2.35 mm long; third tarsomere 2.10 mm long, with stout lateral process, 0.95 mm long; fourth tarsomere indistinctly preserved; ungues incompletely preserved, strongly curved, 1.44 mm long. Mesotrochantora small, c. 2.40 mm long and c. 2.20 mm wide. Mesofemora very slender, somewhat curved, inflated slightly at both the base and geniculae, 11.26 mm long and 1.46 mm wide at midlength. Mesotibiae similar in form to protibiae but somewhat larger and more acutely subovoid in lateral aspect, 10.75 mm long; left mesotibia incompletely preserved in dorsal aspect; right mesotibia preserved in lateral aspect, 3.36 mm wide at midlength, with two short subapical spurs preserved. Left mesothoracic leg (L2 in Fig. 2) with tarsus preserved in dorsal aspect, 8.17 mm long; basitarsus small, 1.15 mm long; second tarsomere 1.25 mm long with broad, incomplete lateral processes at least 1.49 mm long; third tarsomere 1.28 mm long with stout lateral processes at least 0.62 mm long; fourth tarsomere basally inflated, apically slender, 2.18 mm long; ungues at least 2.08 mm long. Right mesothoracic leg (R2 in Fig. 2) with tarsus preserved in lateral aspect, c. 8.10 mm long; basitarsus small, 1.55 mm long; second tarsomere 1.15 mm long with broad, apically incomplete lateral process at least 1.58 mm long; third tarsomere 1.23 mm long with incompletely preserved lateral process at least 0.95 mm long; margin between fourth tarsomere and ungues indistinct, combined length c. 3.91 mm. Metafemora large, robust, 19.78 mm long. Metatibiae markedly shorter than metafemora, 12.58 mm long; acutely subovoid in form, though more elongate than pro- and mesotibiae. Left metatibia preserved in dorsal aspect, somewhat crushed, with poorly preserved spines along dorsolateral margins; first apical spur 3.62 mm long; second apical spur 3.07 mm long; third apical spur incompletely preserved and visible only as a thin, 0.86 mm long fragment immediately adjacent and inferior to the second apical spur; sixth apical spur partially preserved in lateral aspect, 2.88 mm long and 0.96 mm wide apically. Right metatibia preserved in lateral aspect, 3.29 mm wide at midlength, with bases of poorly preserved spines visible along the dorsal margin; first apical spur blade-like, 3.80 mm long; second apical spur incompletely preserved immediately adjacent and inferior to the first apical spur, 2.06 mm long. Left metathoracic leg (L3 in Fig. 2) with tarsus preserved in dorsal aspect, at least 8.76 mm long; basitarsus well-developed, at least 2.80 mm long, with large, broadly paddle-like lateral processes at least 2.41 mm long and 1.78 mm wide; second tarsomere short, 0.98 mm long, with large, blade or paddle-like lateral processes at least 2.28 mm long and 0.94 mm wide; third tarsomere incomplete, at least 1.98 mm long; combined length of fourth tarsomere and ungues c. 2.98 mm. Right metathoracic leg (R3 in Fig. 2) with tarsus preserved in lateral aspect, at least 9.50 mm long; basitarsus well-developed, at least 2.66 mm long, with large, paddle-like lateral process at least 2.37 mm long; second tarsomere short, 1.04 mm long, incompletely preserved; third tarsomere indistinct, with basal part of small lateral process visible; fourth tarsomere and ungues poorly preserved, combined length 3.97 mm.
Comments
The photograph of the holotype provided by Martins-Neto (2007: fig. 1A) is of rather poor quality, though it is obvious that the specimen is not as well preserved as MfNB-I.2079. Moreover, the accompanying drawing (fig. 1B) is not only incomplete (for reasons that are unclear, the drawing only depicts part of the specimen) but does not correspond entirely with features clearly visible in the photograph. Nevertheless, the photograph shows sufficient details for the identification of the holotype as a schizodactylid; namely the presence of distinctive paddle-like lateral processes on the tarsi (clearly visible on both metatarsi though misidentified as ‘well-developed pulvilli’ by Martins-Neto), and apically coiled wings (not mentioned in the original description). Moreover, the holotype is clearly conspecific with MfNB-I.2079, agreeing with it not only in the relative proportions of the legs and overall size and habitus, but also in the distal origin of the metabasitarsal processes.
Schizodactylus groeningae represents the only fossil record of Schizodactylidae and confirms the antiquity of an extant lineage hitherto unknown from the fossil record. Especially significant is that the species belongs to an extant, albeit relict genus, suggesting that the initial radiation of the Schizodactylidae occurred at least during the Jurassic if not earlier. Moreover, the presence of Schizodactylus groeningae represents the only record of Schizodactylidae from the New World, confirming presence of the family in the Atlantic rift zone of South America prior to its complete separation from Africa. An arid or semi-arid local environment for the Crato hinterland was first suggested by Martill (1993) and later supported by the discovery of solifuges (Selden and Shear 1996) and diplurid spiders (Selden et al. 2006). The development of thick and laterally extensive sandstones within the Crato Formation (Heimhofer and Martill 2007) provides direct evidence for the existence of large local sand bodies and the preserved root balls of fossil plants often consist largely of sand-rich palaeosols (Mohr et al. 2007). Evidence for seasonal flash flooding is also well attested (see Martill et al. 2007 and contributions therein) and might explain how the terrestrial elements of the biota were transported into the Crato lagoon. The presence of Schizodactylus in sandy, xeric monsoonal environments today is therefore, entirely consistent with the hypothesised palaeoenvironment and would suggest that the habitat preferences of schizodactylids have changed little in over 100 million years.
Taxon Treatment
- Heads, SW; Leuzinger, L; 2011: On the placement of the Cretaceous orthopteran Brauckmannia groeningae from Brazil, with notes on the relationships of Schizodactylidae (Orthoptera, Ensifera). ZooKeys, 77: 17-30. doi
Other References
- Heimhofer U, Martill DM (2007) The sedimentology and depositional environment of the Crato Formation. In: Martill DM, Bechly G, Loveridge RF (Eds) The Crato Fossil Beds of Brazil, Window into an Ancient World. Cambridge University Press, Cambridge, 44–62.
- Martill DM (1993) Fossils of the Santana and Crato Formations, Brazil. Field Guide to Fossils 5. Palaeontological Association, London, 159 pp.
- Martill DM, Bechly G, Loveridge RF (2007) The Crato Fossil Beds of Brazil, Window into an Ancient World. Cambridge University Press, Cambridge, [xvi] + 625 pp.
- Martins-Neto RG (2007) New Orthoptera Stenopelmatoidea and Hagloidea (Ensifera) from the Santana Formation (Lower Cretaceous, northeast Brazil) with description of new taxa. Gaea 3:3-8.
- Mohr BAR, Bernardes-de-Oliveira MEC, Loveridge RF (2007) The macrophyte flora of the Crato Formation. In: Martill DM, Bechly G, Loveridge RF (Eds) The Crato Fossil Beds of Brazil, Window into an Ancient World. Cambridge University Press, Cambridge, 537–565.
- Selden PA, Costa Casado F, Mesquita MV (2006) Mygalomorph spiders (Araneae: Dipluridae) from the Lower Cretaceous Crato Lagerstätte, Araripe Basin, north-east Brazil. Palaeontology 49:817-826.
- Selden PA, Shear WA (1996) The first Mesozoic solifuge (Arachnida) from the Cretaceous of Brazil, and a redescription of the Palaeozoic solifuges. Palaeontology 39:583-604.
Images
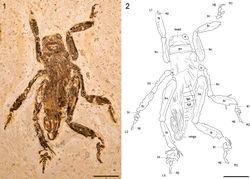 Figures 1-2. Schizodactylus groeningae (Martins-Neto, 2007), comb. n. from the Lower Cretaceous Crato Formation of Brazil. 1 Photograph of MfNB-I.2079 2 Camera lucida drawing of MfNB-I.2079. Abbreviations: R1 – right prothoracic leg; L1 – left prothoracic leg; R2 – right mesothoracic leg; L2 – left mesothoracic leg; R3 – right metathoracic leg; L3 – left metathoracic leg; bt – basitarsus; fm – femur; pn – pronotum; tb – tibia; tr – trochanter; ug – ungues. Scale bars represent 10 mm.
Figures 1-2. Schizodactylus groeningae (Martins-Neto, 2007), comb. n. from the Lower Cretaceous Crato Formation of Brazil. 1 Photograph of MfNB-I.2079 2 Camera lucida drawing of MfNB-I.2079. Abbreviations: R1 – right prothoracic leg; L1 – left prothoracic leg; R2 – right mesothoracic leg; L2 – left mesothoracic leg; R3 – right metathoracic leg; L3 – left metathoracic leg; bt – basitarsus; fm – femur; pn – pronotum; tb – tibia; tr – trochanter; ug – ungues. Scale bars represent 10 mm.
